1,400 words
Sign up for the Blank Horizons mailing list here.
Header image taken from here.
As we saw in Part I, causation is at the core of Integrated Information Theory (IIT). One of the postulates of IIT is that consciousness contains information, which is measured as the extent to which irreducible mechanisms in the physical substrate of consciousness (PSC) constrain its cause-effect structure. This structure is the set of probability distributions of all possible past and future states of each mechanism. In other words, if current states of the system are associated with a very narrow range of past and future states, then the system is more conscious, so long as those states are also maximally irreducible.
IIT always associates causes with past states of the system at hand, and effects with future states. Thus, IIT’s notion of causation aligns with the conventional definition; namely, causation is linear, such that causes always precede effects.
There are two reasons to think that the physical substrate of consciousness – that is, the brain – does not obey linear causation. First, as the neuroscientist György Buzsáki notes in his book Rhythms of the Brain, causes of brain phenomena are often simultaneous with their effects. According to Buzsáki, “in an oscillatory [neuronal] system, most or all neurons with reciprocal, one-way connections or no direct connections may discharge with a zero time lag (i.e., simultaneously), making linear causation impossible.” (Buzsáki argues that oscillations underlie many important cognitive and perceptual phenomena, including consciousness.)
Secondly, Buzsáki also claims that the brain exhibits circular causation due to its nonlinear self-organization. A nonlinear system, roughly speaking, is one in which the whole is greater than the sum of its parts. (This definition actually aligns with Tononi’s notion of irreducibility, which, he claims, explains the unity of consciousness.) This nonlinearity is attributable to a bidirectional relationship between the whole and its parts; local interactions produce global patterns, as they do in all systems, but then the global patterns constrains the degrees of freedom that govern the local activity. To quote Buzsáki, “While the parts do cause the behavior of the whole, the behavior of the whole also constrains the behavior of its parts according to a majority rule; it is a case of circular causation. Crucially, the cause is not one or the other but is embedded in the configuration of relations. In fact … in [nonlinear, self-organizing] systems the cause is always circular.”
In both of these cases, it becomes difficult to distinguish between causes and effects, so the quantity of integrated information in the brain is ambiguous. Arguably, the feedback loop in the latter case can be decomposed into linear causation; even if local and global phenomena form a cycle, this cycle nevertheless produces a linear succession of events: local –> global –> local, etc. Hence, any event that occurs in the present will still have a definite cause and effect. However, calculating integrated information requires very precise knowledge of the input mechanisms that are affecting the brain, that is, which neurons or groups of neurons are activated in the brain. But global, emergent patterns are precisely the kinds of phenomena that cannot be traced to individual mechanisms in the brain, even if these are higher-order mechanisms. As Buzsáki notes, these global patterns are order parameters; these include, for instance, variance in the synchrony of networks, that is, the tendency of neurons to synchronize with the mean signal of a network. There is no direct method, as far as I am aware, of relating this order parameter to the firing rates of neurons, yet Tononi has suggested that the latter quantity serves as the mechanism that governs the degree of integrated information in the brain.
In this part of the essay series, I will address the latter objection to the claim that consciousness obeys linear causation; in particular, I will propose an alternative method of determining “causation” by measuring the reciprocal relationships between the local and global scales of the brain.
Causation: a measure of the extent to which the local and the global co-determine each other
If the order parameter of consciousness is variance in the synchrony of brain networks, then the local interactions that modulate the order parameter are changes in the excitation and inhibition of neurons. (Incidentally, neurons trigger action potentials, the unit of information in the brain, when the difference between the sum of excitatory and of inhibitory inputs exceeds a certain threshold.) The excitation-inhibition balance of the network then determines the oscillations that occur at the global scale, i.e. at the network level; the wave goes up during the excitatory phase and down during the inhibitory phase. When neurons tend to synchronize their firing with the network’s oscillations, then variance and hence the order parameter is low; if neurons activate out-of-step with the network, however, the order parameter is high.
However, certain neurons in the network have a larger effect on the mean oscillation of the network than others. Indeed, some may have disproportionately more synapses, or connections, with other neurons. Therefore, we can reduce our uncertainty about the global state to a greater degree by determining the states of these important, or informative, neurons than by doing so for the unimportant neurons. In other words, the informative neurons constrain the repertoire of global activity in the network to a larger extent than less significant neurons.
Geometrically, this statement implies that informative neurons specify a more constrained region (or “submanifold”) of the order parameter space (Fig 1). This space is the set of all possible order parameter values. If there are n networks in the brain that are relevant for consciousness, then there are n dimensions (or coordinates) that are needed to identify a point in the order parameter space. In 3 dimensions, the order parameter space would be a sphere. One neuron can be a part of many networks, so t the activity of one neuron is not necessarily associated with a region in just one dimension of the order parameter space. In the first step of computing causation, we label a neuron as informative if it encodes a submanifold that is sufficiently constrained.
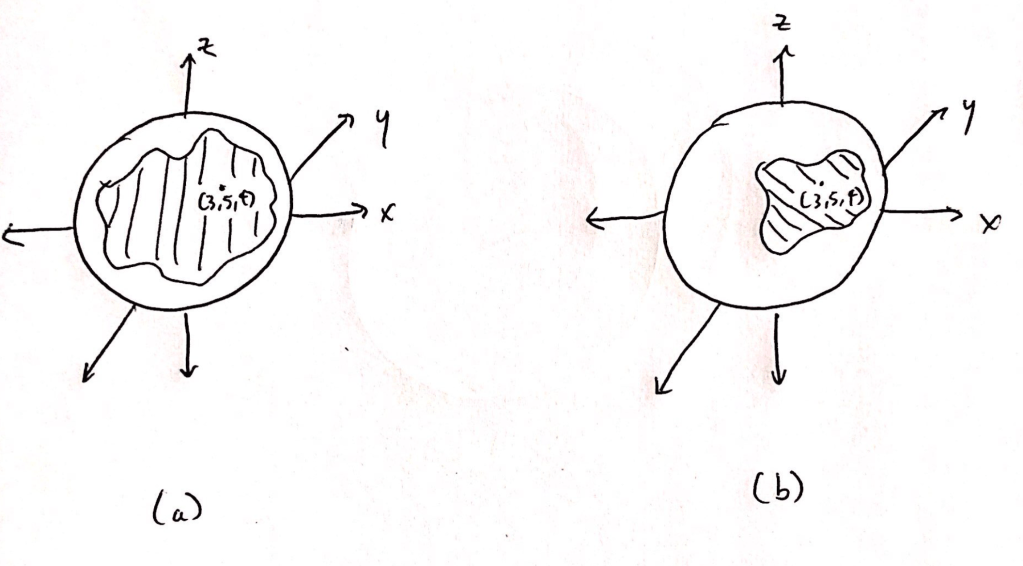
In the second step, we quantify the lengths of the possible paths within the submanifolds specified by the informative neurons. A path is simply a trajectory from one point to another on connected segments of the submanifold; if the submanifold is not connected, that is, if there is an unbridgeable gap between any two parts, then there are no paths between the unconnected segments (Fig 2). The brain has more causal power if the paths within the submanifold tend to be very long. Then, the network is capable of producing large changes in the order parameter of consciousness.
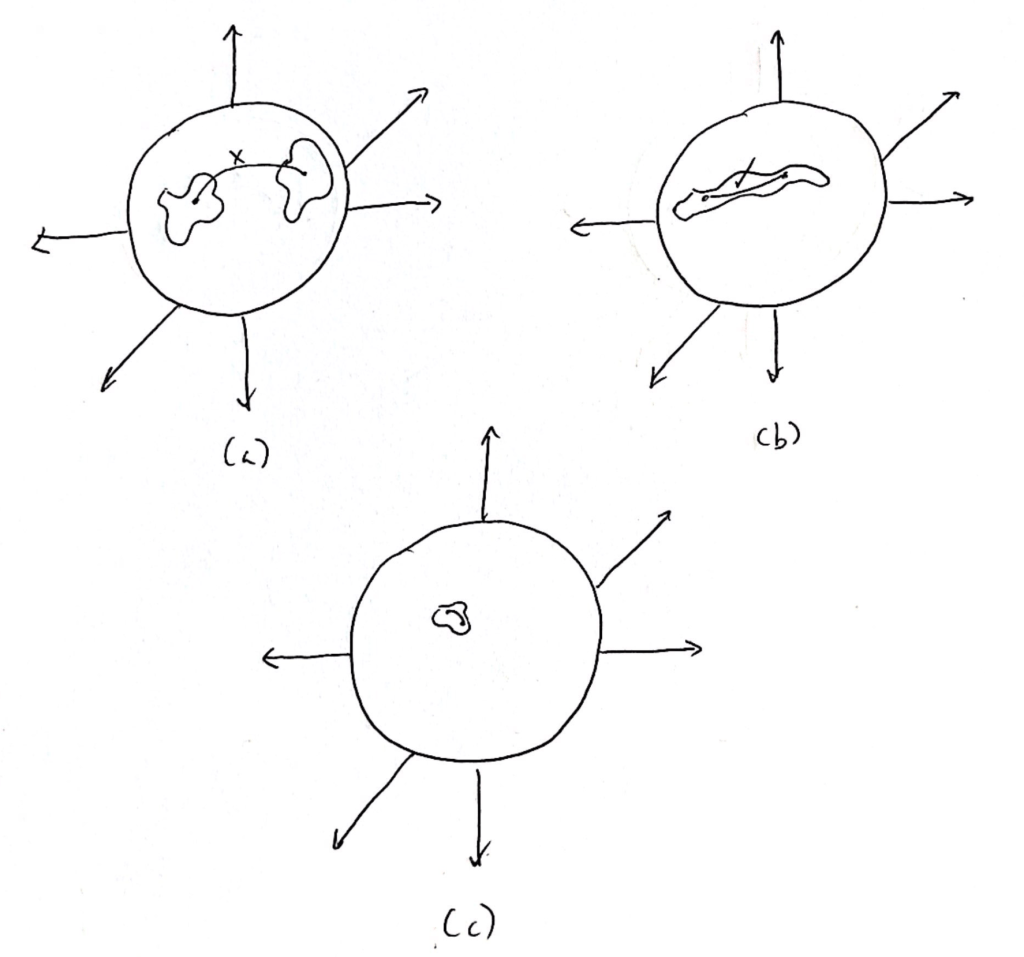
How does this method of quantifying causation capture the reciprocal interactions between local and global events in the brain? Large changes in the (global) order parameter have a greater effect on the local excitation and inhibition of neurons. If the order parameter jumps from a high value to a low value from one moment to the next. then individual neurons will start synchronizing with the global oscillations of the network, thereby changing their own firing patterns. Thus, the brain exhibits greater causal power if it exerts more influence over the order parameter, that is, if the submanifolds of the informative neurons have longer paths. The order parameter modulates the local interactions, and then the local events decide the value of the order parameter; local and global phenomena co-determine each other.
